
Авторы: Michael D. Simone-Finstrom и Marla Spivak, 2012 год.
Оригинал статьи на английском языке
Введение
Постоянное давление, оказываемое паразитами, привело к тому, что все виды животных развили множество механизмов, способных противостоять инфекциям. У беспозвоночных видов основными способами защиты от паразитов являются индивидуальные барьеры и физиологические виды защиты. Однако некоторые виды поведения и другие неиммунологические защитные механизмы также могут эффективно снижать интенсивность передачи паразитов и интенсивность инфицирования. У социальных насекомых поведение, снижающее нагрузку паразитов на уровне всей колонии, называется «социальным иммунитетом». Одним из примеров поведенческой защиты является использование растительных смол. Медоносные пчёлы собирают растительные смолы и включают их в архитектуру своего гнезда. Такое использование смол может снизить регулярную нагрузку на иммунитет отдельной пчелы. Поскольку высокая активация каждого индивидуального иммунитета может привести к повышенным энергозатратам на уровне всей пчелосемьи, сбор смол может принести пользу как индивидууму, так и всей семье. Однако использование смол в качестве прямой защиты от патогенов остается не до конца изученным. Здесь мы приводим доказательства того, что семьи медоносных пчёл могут самостоятельно лечить себя растительными смолами в ответ на грибковую инфекцию. Самолечение обычно определяется как индивидуальное реагирование на инфекцию путем приема в пищу или накопления непищевых компонентов или растительных материалов. Наши результаты показывают, что пчелосемьи увеличивают объем добычи смол (и выработки прополиса), будучи подвержены грибковому паразиту (Ascophaera apis: аскосфероз, или известковый расплод). Кроме того, пчелосемьи, экспериментально обогащенные смолой, снижали интенсивность заражения этим грибковым паразитом. Если рассматривать самолечение, то это особенно уникальный пример, потому что оно действует на уровне всей пчелосемьи. Большинство случаев самолечения связано с фармакофагией, когда организм меняет свой рацион питания в ответ на прямое заражение паразитом. В нашем случае с медоносными пчелами смолы не употребляются в пищу, а используются внутри улья взрослыми пчелами, подвергшимися воздействию грибковых спор. Таким образом, пчелосемья, являясь единицей отбора, может реагировать на инфекцию путем самолечения, увеличивая число особей, которые добывают смолу.
Вступление
Организмы развили множество защитных механизмов, чтобы переносить паразитарную инфекцию или противостоять ей [1], [2]. Под паразитами мы понимаем и макропаразитов, таких, как членистоногие, и микропаразитов, таких, как бактерии и грибы, которые живут на хозяине или в нем и ухудшают его общее физическое состояние. Индивидуальные барьеры и физиологические механизмы, такие как кутикулы (Кутикула – внешний слой кожи насекомых, не имеющий клеточного строения и являющийся продуктом выделения гиподерм) и индуцибельные антимикробные пептиды у насекомых, являются общими способами борьбы с инфекцией; однако организмы также используют различные поведенческие черты, чтобы бороться с паразитами [2], [3]. Социальные виды (к которому относятся и пчёлы) добавляют еще один уровень сложности в свои защитные механизмы, у них защита функционирует как на индивидуальном, так и на групповом уровнях. У эусоциальных насекомых (например, медоносных пчёл и муравьев) можно наблюдать как физиологические, так и поведенческие виды защиты, которые имеют последствия для здоровья как отдельных особей, так и всей колонии. Социальный иммунитет — это явление, при котором поведение индивида снижает паразитарную нагрузку и паразитарный стресс на групповом уровне [4]. Например, было показано, что включение растительных смол внутрь гнезда медоносной пчелы (Apis mellifera) снижает бактериальную нагрузку семьи и снижает общие «расходы» на индивидуальную иммунную функцию, что может положительно повлиять на состояние семьи. [5], [6]. Представленные здесь исследования свидетельствуют о том, что использование смол медоносными пчелами может являться примером механизма самолечения на уровне всей пчелосемьи, что поддерживает концепцию о том, что сбор смол у медоносных пчел является одной из форм социального иммунитета. Чтобы действительно классифицировать какой-то признак как самолечение у животных, он должен быть адаптивно пластичным, то есть индивид (или колония в данном случае) должен демонстрировать данное поведение с более высокими показателями при наличии паразитов, и с более низкими показателями или вообще не выполнять его при здоровом состоянии [7].
Наиболее хорошо изученные примеры самолечения включают глотание цельных листьев особями различных видов приматов для устранения инфекций нематод [8] — [11], или поедание вторичных метаболитов растений, особенно различными гусеницами и шмелями [7], [12]–[14]. Эти примеры описывают поведение, которое подпадает под термин фармакофагия, то есть прием непитательных веществ для целей, отличных от энергетических потребностей [15]. (Тут надо заметить, что в русском языке термин фармакофагия имеет скорее отрицательную коннотацию — чрезмерное, часто болезненное употребление лекарств, в том числе без назначения врача. В оригинальной статье акцент другой.)
Другие виды самолечения включают использование зеленой массы и вторичных метаболитов растений наружно, а не через прием внутрь. Фармакофория определяет такие виды поведения, при которых растительные материалы собираются и используются вовне [16] (например, при строительстве гнезд или уходе за ними [5], [6], [17]–[21]). В случае фармакофории или фармакофагии поведение может быть конститутивно выраженным и, следовательно, профилактическим, либо условно выраженным и, следовательно, формой самолечения. Например, в ряде исследований было установлено, что ароматические листья, используемые при постройке гнезд европейскими скворцами и голубыми синицами, могут снижать паразитарную нагрузку и положительно влиять на иммунокомпетентность птенцов [18], [21]. Однако, эти птицы не увеличивают сбор таких листьев ответ на высокие нагрузки паразитов, поэтому это, с большой вероятностью можно отнести к профилактическому поведению. Другой пример использования смол был проиллюстрирован в серии лабораторных исследований социального швейцарского древесного муравья (Formica paralugubris). Использование смолы в гнезде является одной из форм социального иммунитета, поскольку она снижает общую микробную нагрузку внутри колонии [19] и может повысить выживаемость зараженных паразитами особей [22], [23]. Данное поведение является ярким примером фармакофории—муравьи используют смолу профилактически, чтобы принести пользу колонии. При этом было обнаружено, что оно не является примером самолечения, поскольку индивидуумы не увеличивают сбор смолы, когда подвергаются усиленной атаке паразита [24].
Медоносные пчелы собирают смолы из различных видов растений по всему миру. В умеренных регионах принято считать, что основными источниками являются Populus spp, а в тропических регионах широко используются травянистые кустарники (например, Baccharis dracunculifolia) [6], [25]–[27] и смолообразующие цветочные виды (например, Clusia spp.). «Прополис» (по-гречески «про» – перед, или оборона, а «полис» – город) — это пчеловодческий термин для обозначения смол, собираемых и используемых в улье медоносными пчелами. Неуправляемые, «лесные, дикие» пчелиные семьи, обычно живущие в древесных дуплах и полостях, выстилают всю внутреннюю поверхность улья тонким слоем смолы, смешанной с различными количествами воска, создавая так называемую «прополисную оболочку» [6], [28].
Наши предыдущие исследования показали, что медоносные пчелы в обогащенном смолой улье способны снижать индивидуальные энергозатраты на иммунную функцию за счет общего снижения бактериальной нагрузки семьи [5]. Как и у швейцарских древесных муравьев [19], использование смолы медоносными пчелами является профилактической фармакофорией и функционирует как тип социального иммунитета [4], в результате чего включение смолы в гнездо отдельными медоносными пчелами приносит пользу иммунитету на уровне семьи.
Здесь мы идем еще дальше, спрашивая, могут ли семьи медоносных пчел заниматься самолечением, собирая растительные смолы в дополнение к их профилактическому использованию. Мы следили за показателями сбора кормов семьями до и после воздействия на семью микробных паразитов. Это исследование проводилось с начала июля по начало сентября 2008, 2009 и 2010 годов. Во все три года колонии подвергались воздействию грибкового паразита Ascosphaera apis, возбудителя личиночной болезни аскосфероза, или «известкового расплода» (СВ). В 2009 году некоторые семьи были дополнительно заражены бактериальным паразитом Paenibacillus larvae (личинка возбудителя болезни Американский гнилец, American foulbrood, или AFB) или подверглись воздействию спор почвенного гриба-энтомопатогена Metarhizium anisopliae. Поскольку Метаризий не является патогенным для медоносных пчел, его использовали в качестве контроля за повышенными микробными нагрузками в неблагополучных колониях.
Результаты
В целом, в семьях с заражением аскосферозом увеличился сбор смолы (рис. 1А-Д). Все показатели приводятся с допустимыми отклонениями. В течение трех лет в семьях, пораженных аскосферозом, среднее число пчёл, несших в гнездо смолу (смоляных фуражиров) составляло 6,0±1,0 вне заражения, и 8,7±0,9 после заражения за 15-минутный период наблюдения. В незараженных колониях в среднем было 9,1±1,4 смоляных фуражиров до заражения и 8,2±1,5 после заражения. Средняя разница в общем количестве смоляных фуражиров (сумма «после» минус сумма «до» заражения) составила 2,8±1,1 для колоний с аскосферозом и -0,9±1,2. Общее количество смоляных фуражиров неизменно составляло меньшинство от общего количества фуражиров (рабочих пчел, несущих корм в гнездо) (≈1% или менее), поэтому это кажущееся незначительным изменение среднего числа пчел, добывающих смолу в течение 15-минутного интервала, является биологически значимым.
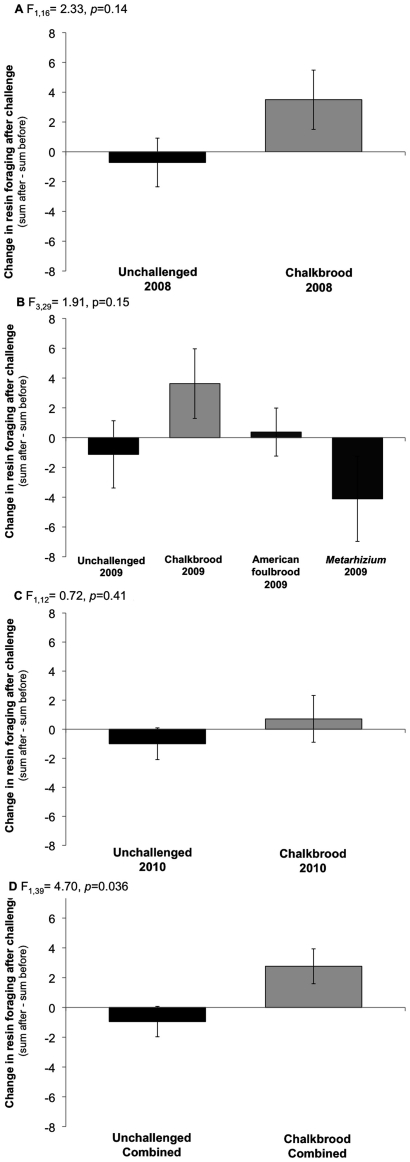
По результатам двустороннего ANOVA-анализа комбинированных данных, рассматривающего воздействие аскосфероза и год наблюдения в качестве основных факторов, удалось определить, что наблюдалось значительное увеличение скорости заготовки смолы при наличии заражения аскосферозом (F1, 46 = 4,70, p = 0,036; рис. 1D). Не было замечено разницы в зависимости от года наблюдения (F2, 46 = 0,33, p = 0,72) а также взаимозависимости между годом и аскосферозом (F2,46 = 0,30, p = 0,74). Независимый анализ по годам показал, что в 2008 году наблюдалось умеренное, но незначительное увеличение заготовки смол после заражения аскосферозом (F1, 16 = 2,33, p = 0,14; рис. 1А). В 2009 году колонии подверглись воздействию двух других патогенов. Полный модельный анализ показывает, что заготовка смол в семьях с аскосферозом возросла значительно по сравнению с колониями с метаризием Metarhizium (Р = 0,025) и умеренно по сравнению с незараженными колониями (Р = 0,17). Изменение общего числа смоляных фуражиров, наблюдаемое для колоний с поражением американским гнильцом, несущественно отличалось от других поражений (Р>0,18; см. рис. 1Б). В 2010 году наблюдалось незначительное увеличение темпов заготовки смолы после заражения аскосферозом. (F1, 12 = 0,72,Р = 0,41; Рис.1). 1С).
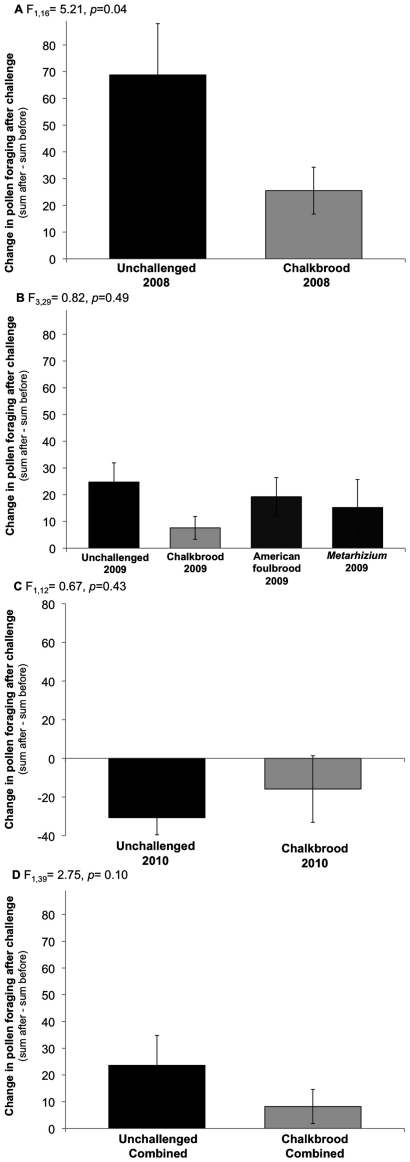
Увеличение темпов заготовки смол после заражения не было результатом увеличения общих темпов заготовки кормов в семьях с аскосферозом, о чем свидетельствуют подсчеты пыльцевых фуражиров (пчел, приносящих пыльцу). (Рис.1). 2А-Д). Собиратели пыльцы, как и следовало ожидать, были очень многочисленны, особенно по сравнению с собирателями смолы. В среднем за три года у семей с аскосферозом наблюдалось 50,3±5,3 пыльцевых фуражиров до заражения и 58,5±4,7 после заражения за 3 мин наблюдения (или за 15 мин для смоляных фуражиров).
Напротив, средние значения (+SE) для незараженных колоний составили 54,6±6,5 до заражения и 78,2±8,0 после него. Это привело к тому, что средние общие различия (сумма «после» минус сумма «до») в общем количестве собирателей пыльцы между периодами «до» и «после» заражения составили 8,2±6,2 для семей с аскосферозом и 23,6±6,7 для незараженных семей. Двустороннее ANOVA-моделирование по виду заражения и году наблюдения как основным факторам для комбинированных данных показало, что пораженность аскосферозом не оказала существенного влияния на изменение заготовок пыльцы до и после заражения (F1, 46 = 2,75, p = 0,10; рис. 2.Д). Эффект года сказался наибольшим увеличением сбора пыльцы независимо от аскосфероза — в 2008 году, затем аналогично в 2009 году, а в 2010 году наблюдалось незначительное снижение (F2,46 = 19,02, p<0,0001). Анализ отдельных лет показал, что в 2008 году у незараженных семей наблюдалось существенное увеличение добычи пыльцы после заражения по сравнению с колониями с уже имеющимся аскосферозом (F1, 16 = 5.21, p = 0.04; рис. 2А). Существенной разницы наличие заражения в 2009 году не показало (F3,29 = 0,82, p = 0,49; рис. 2B) также и в 2010 году (F1,12 = 0,67, p = 0,43; рис. 2С).
Уровень заражения аскосферозом в семьях 2008.
| Семья | Заражение | Известкованные личинки | Известкованные личинки | Всего | |
| Воздействие | Учет 1 | Учет 2 | |||
| Мало смол | Незараженные | 0 | 0 | 0 | A |
| Аскосфероз | 42.3±25.1 | 65.8±38.2 | 108.2±49.0 | B | |
| Много смол | Незараженные | 3.2±1.6 | 2.2±1.4 | 5.3±1.7 | A |
| Аскосфероз | 1±0.5 | 13.7±7.2 | 14.7±7.5 | B |
Эти данные описывают все семьи, наблюдаемые в 2008 году, независимо от ранжирования по сбору смол (n = 5 смол мало, семья не заражена; n = 6 одинаково для смол мало, семья заражена, смол много, семья незаражена и смол много, заражена).
Богатые смолой незараженные семьи, положительные по аскосферозу, либо имели стойкие низкие уровни инфекции (1 семья), либо низкие уровни инфекции только в один момент времени (2 семьи). Буквы указывают на значительную разницу в общем количестве известкованного расплода, основанных на двусторонней ANOVA.
Обсуждение
Исследования показали, что интенсивность сбора смол увеличивалась, когда свободно живущие пчелиные семьи подвергались воздействию грибкового агента, вызывающего заболевание личинок, аскосфероза (A. apis), что позволяет предположить, что пчелиные семьи могут заниматься самолечением в ответ на этот конкретный патоген. Представленные здесь данные также свидетельствуют о том, что помимо косвенного иммунного эффекта, смолы могут напрямую воздействовать на некоторые специфические грибковые паразитарные заражения. Хотя есть предварительные доказательства того, что экстракты прополиса эффективны против различных грибковых паразитов, практических знаний об активности прополиса против аскосфероза пока недостаточно [6]
Результаты, представленные здесь в рамках исследования 2008 года, свидетельствуют о том, что богатая смолами среда может напрямую влиять на снижение интенсивности аскосферозной инфекции и оказывать ингибирующее воздействие на рост этого гриба. В свете этого сбор смол пчелиными семьями может быть новым примером самолечения, когда индивидуальное проявление поведенческой черты изменяется под воздействием грибкового патогена. Медоносные пчелы не поглощают смолу, а аскосфероз паразитирует на личинках, а не взрослых особях — таким образом, сбор смолы взрослыми особями влияет на здоровье всей семьи, т.е. социальный иммунитет. Если пчелы-фуражиры действительно реагируют на некоторых паразитов, увеличивая сбор антимикробных смол, то это особенно уникальный пример самолечения. Поскольку у высоко эусоциальных насекомых (например, медоносных пчел, муравьев, термитов) семья или колония является репродуктивной единицей и фокусом отбора [29], [30], колония может в этом смысле рассматриваться как «индивид».
Заготовка смол встречается относительно редко, особенно у пчёл европейского происхождения. «Окультуривание» медоносных пчел привело к сокращению сбора смол [31], вероятно, потому, что пчеловоды вели селекцию «против», поскольку наличие большого количества липкого прополиса часто затрудняет открытие ульев. Количество прополиса, выстилающего естественную полость гнезда, такое как прополисная оболочка в дупле дерева [6], не было количественно исследовано, но мы обнаружили, что создание прополисной оболочки, содержащей 60 г смол, положительно влияет на иммунитет колонии [5]. Таким образом, увеличение количества смоляных фуражиров (даже относительно незначительное увеличение, как видно из представленного исследования), вероятно, имеет большой биологический эффект. Несмотря на то, что мы в объединенном трехлетнем исследовании обнаружили статистически значимое влияние заражения аскосферозом на интенсивность заготовки смол, тенденции в каждом году предполагают, что увеличение общей интенсивности заготовок в течение одного года даст тот же результат.
В 2010 году, возможно, интенсивность заготовки кормов была в целом низкой, о чем свидетельствует общее снижение добычи пыльцы после заражения, поэтому даже скромное увеличение добычи смол, которое мы обнаружили, говорит о важности такого поведения на уровне пчелосемьи.
Большинство других случаев самолечения, наблюдаемых у позвоночных и беспозвоночных животных, связаны с изменением их рациона питания (фармакофагия) в ответ на прямое заражение паразитом [7], [10]. В случае с медоносными пчелами смолы используются внутри гнезда (фармакофора) и не употребляются в пищу. [16]. Кроме того, именно семья как целое реагирует на паразитарную инфекцию, а не отдельные особи. Это важная разница в наших исследованиях между медоносными пчелами и другими очевидными примерами самолечения: шимпанзе с активными нематодными инфекциями глотают целиком листья определенных растений[8], [9], [32]; зараженные паразитоидами гусеницы G. incorrupta поглощают непитательные алкалоиды [7]; гусеницы Spodoptera littoralis предпочтитают потреблять высокобелковую еду при заражении вирусом [12]. В последних двух примерах с одиночными насекомыми прием этих веществ приводит к большим затратам на приспособление (например, сокращение продолжительности жизни), когда особи не инфицированы. Хотя пчелы не потребляют смолы в пищу, поиск смолы, вероятно, является дорогостоящим процессом, потому что как поиск смолы, так и ее обработка в улье занимает много времени и не дает очевидного прямого пищевого вознаграждения, как поиск нектара или пыльцы. Однако сбор смолы действительно функционирует как механизм социального иммунитета [6]. Включение смол в среду гнезда снижает общую бактериальную нагрузку в семье, либо за счет ингибирования вследствие прямого контакта, либо за счет высвобождения летучих соединений [33], и поэтому позволяет особям меньше инвестировать в иммунную функцию [5]. Поскольку сильная активация индивидуального иммунитета может иметь издержки на уровне всей семьи [34], то все способы, снижающие необходимость повышенного иммунного ответа каждого индивида, могут быть полезны для продуктивности колонии. Поэтому различные затраты индивидуума могут быть компенсированы преимуществами использования смол в семье, поскольку жизнеспособность индивида в значительной степени определяется успехом всей семьи медоносных пчел.
Поскольку только личинки могут заразиться аскосферозом, поначалу было удивительно, что взрослые особи изменили свое поведение в ответ на повышенный уровень паразита, который непосредственно не влияет на них самих. Однако с точки зрения социального иммунитета семья является зараженной единицей, и поэтому семья отреагировала на это увеличением числа смоляных фуражиров. Наш вывод о том, что уровень сбора смолы изменился только после заражения аскосферозом, а не гнильцом или грибом Metarhizium, требует дальнейшего изучения. Чтобы создать заражение аскосферозом, мы гомогенизировали споры в смеси пыльцы, таким образом взрослые пчелы обрабатывали и, возможно, поглощали их. Споры не прорастают в кишечнике взрослой пчелы, но могут оставаться там, и взрослые особи являются главным распространителем спор аскоферы по всей колонии через личиночное питание [35] — [37]. Таким образом, отдельные взрослые пчелы подвергались общему процессу увеличения грибковых спор по всей колонии в результате заражения аскосферозом, хотя семьи в основном проявляли легкие симптомы заболевания или не имели клинических симптомов. Отсутствие подобной реакции в семьях, обработанных энтомопатогеном Metarhizium, может быть связано с тем, что медоносные пчелы обычно не подвергаются воздействию этого типа почвенного гриба.
Медоносные пчелы обычно удаляют мусор с пола гнезда [38], где в основном и находился Метаризиум в заражаемых семьях. Поскольку этот гриб не размножался и не накапливался в улье, вполне вероятно, что семьи, зараженные Метаризием просто удаляли мусор со спорами из гнезда. Однако исследования, проведенные по использованию смолы муравьями Formica paralugubris после заражения Метархизием, который естественным образом поражает этот вид, также показали, что эти муравьи не увеличивают сбор под воздействием этого паразита [24], хотя присутствие смолы может помочь снизить смертность. Таким образом, вполне возможно, что этот тип самолечения является более специфической реакцией у медоносных пчел и, возможно, других видов, хотя на этом фронте необходимо провести дополнительные исследования.
Неясно, почему семьи не увеличили сбор смол в ответ на вызов бактерий американского гнильца AFB. Видимо, набор физиологических защитных механизмов, особенно антимикробные пептиды, у медоносных пчел, по-видимому, больше ориентирован на контроль бактериальных паразитов [41], в то время как клеточные иммунные механизмы (например, клеточная инкапсуляция), вероятно, участвуют в индивидуальной защите от грибковых паразитов [39], [40]. Однако в лабораторных условиях было обнаружено, что экстракты прополиса действуют против гнильца в лабораторных культурах [42] а также пчелосемьях в природе, которых подкармливали экстрактами прополиса в сахарном сиропе [43]. Было также показано, что богатая смолой среда снижает общую бактериальную нагрузку в семьях [5]. Таким образом, можно предположить, что самолечение смолой против бактериальной инфекции, в дополнение к грибковой инфекции, было адаптивной реакцией и требует дальнейшего изучения. Другой аспект, который может влиять на реакцию на паразитов с помощью одних защитных механизмов, а не других, заключается просто в том, что существует ряд защитных механизмов, которые отдельные особи и семьи предпочитают использовать против одного паразита (например, гигиеническое поведение, сбор смолы, уход за расплодом, физиологические меры [2]). Учитывая это, в настоящее время неизвестно, как эти средства защиты используются в пределах одной семьи и как они используются совместно.
В представленном здесь исследовании, а также в исследовании, проделанном с гусеницей G. incorrupta [7], наличие паразитов увеличивает скорость выполнения рутинного поведения вместо инициации атипичного поведения (например, заглатывание листьев у приматов [8]-[11]). Поведенческий механизм, инициирующий сбор смол медоносной пчелой в ответ на заражение семьи конкретным грибковым паразитом, в настоящее время неизвестен. В случаях самолечения у позвоночных животных обычно происходит ассоциативное или социальное обучение [11], [44], [45].
Однако те немногие случаи с насекомыми, которые демонстрируют самолечение, не обязательно связаны с обучением, а скорее являются реакцией на изменение гомеостаза организма. У G. incorrupta инфекция вызывает изменения в периферической нервной системе и усиливает активность вкусовых рецепторов к пирролизидиновым алкалоидам (па), что приводит к увеличению потребления богатых ПА источников пищи [46]. Можно предположить, что пчелы повышают сбор смол в ответ на разные степени поражения аскосферозом, руководствуясь обонятельными стимулами. Личинки выделяют специфические химические соединения в ответ на инфицирование аскосферозом, и эти соединения запускают гигиеническое поведение, тип социального иммунитета, при котором пчелы удаляют больных личинок и куколок из гнезда [47].
Эти соединения высвобождаются ранее четкого визуального развития клинических симптомов [47]. Смоляные фуражиры могут также воспринимать сигналы, связанные непосредственно с уровнем микробной среды в семье. Поскольку дикие семьи выстилают всю внутреннюю часть гнезда смолами до и во время строительства сотовых языков, вполне возможно, что бактерии и грибы, обычно встречающиеся в дупле или тканях дерева, также могут индуцировать это поведение. Вероятно, существует много других, не исключающих друг друга стимулов, участвующих активации поведения.[48]. Например, поскольку это самолечение на уровне колонии, то необходимо также учитывать социальные стимулы, например, было отмечено, что танец смоляных фуражиров можно рассматривать как как возможный механизм привлечения других пчел к заготовке смол [26], [49].
До сих пор существует множество вопросов, касающихся сбора и использования смол медоносными пчелами, а также использования смол в животном мире в целом. Его роль как механизма социального иммунитета у пчел и муравьев, вероятно, довольно сложна, включая как прямое воздействие на паразитов, так и более опосредованное воздействие на индивидуальный иммунитет. Выделение смол и вторичных метаболитов растений, по-видимому, является относительно широко распространенной чертой, и многие виды могут использовать эти защитные механизмы растений в качестве механизма защиты от различных паразитов и хищников. Хотя у нас есть некоторые доказательства того, что сбор смолы медоносными пчелами может быть новым случаем фармакофорического самолечения у насекомого, возможно, что это явление более широко распространено, чем считалось ранее.
Материалы и методы
Подготовка пчелосемей
Семьи в 2008 году были подобраны по размеру (~8000 взрослых пчел) и содержались в новых, одинарных стандартных пчеловодческих ульях Лангстрота Langstroth на 9 рамок. На протяжении всего эксперимента семьи содержались в однокорпусном улье. Семьи были разделены между двумя пасеками в Юго-Восточной Миннесоте. Двенадцать ульев были обогащены смолами, путем нанесения на внутренние стенки приблизительно 92,5 г экстракта прополиса в 70% EtOH, чтобы имитировать прополисную оболочку, наблюдаемую в диких пчелиных гнездах [5]. Одиннадцать семей не обогащались смолами, в их ульях было нанесение того же объема 70% EtOH.
В 2009 и 2010 годах семьи были поселены в нуклеусы на четыре рамки и равным количеством взрослых пчел и естественно выведенными сестринскими матками. Все семьи содержались на одной пасеке.
Заражение паразитами
Семьи были подвергнуты заражению грибковым патогеном Ascosphaera apis путем гомогенизации свежих мумифицированных личинок известкового расплода (CB) (т. е. мертвых личинок пчел, инфильтрированных мицелием), и смешивания их с заменителем пыльцы и 50%-ным раствором сахарозы (2008) или со смесью пыльцы и 50% — ным раствором сахарозы (2009, 2010), модифицированной по Гиллиаму и др. [35]. (Назовём из пыльцевыми пирожками, прем. переводчика)
В 2008 году 12 семьям было дано примерно 3,3 эквивалента мумий в 450 г пыльцевых пирожков. В 2009 и 2010 годах 9 и 7 семьям, соответственно, давали 10 эквивалентов личинок в 75 г пыльцевых пирожков. Контрольным семьям давали пыльцевые пирожки без патогена. Уровень аскосферозной инфекции определяли путем подсчета количества мумий (известкованных личинок), присутствующих в расплодных рамках в каждой семье в середине и в конце экспериментального периода. В 2009 году дополнительно девять семей были подвергнуты каждому из трех следующих методов паразитарного воздействия, и девять незараженных семей были использованы в качестве контроля. Для исследования заражения американским гнильцом в 9 семей был введен квадратный участок сота размером 7,5 см из семьи, зараженной американским гнильцом (AFB, Paenibacillus larvae), который содержал личиночные чешуйки AFB [50]. Для заражения Metarhizium anisopliae (энтомопатогенный гриб, который не заражает медоносных пчел) внутренние полы оставшихся 9 семей были посыпаны 75 г порошка M. anisopliae ECS1, содержащего примерно 1×1010 конидий на грамм. Было показано, что это количество не оказывает отрицательного влияния на развитие или здоровье семей [51], и оно использовалось в качестве контроля для увеличения микробной нагрузки в зараженных семьях. Все процедуры были проведены дважды в течение двухнедельного периода испытаний. Уровни инфицирования в результате заражения были измерены один раз в середине и в конце эксперимента.
Сбор смол
Количество собирателей смолы и пыльцы, возвращающихся в улей в период до заражения (июль) и после заражения (август), определялось путем закрытия летка между 12:00 и 16:00 часами каждый день в течение четырех (2008) или шести (2009, 2010) дней в пределах двух недель. Затем подсчитывалось количество пчел со смолой (через 15 мин после закрытия летка) или пыльцой (через 3 мин после закрытия летка) в их корбикулах (корзинках на задних лапках). Поскольку смоляные фуражиры встречаются относительно редко (≈1% от всех фуражиров), 15-минутный период наблюдения был признан достаточным для сбора адекватных данных о количестве смоляных фуражиров и недопущения перегрева закрытых семей. Собиратели пыльцы использовались как показатель общей интенсивности сбора кормов, поскольку они явно занимались добычей пищи; другие пчелы, летающие в улей и из него, могли быть сторожевыми пчелами или разведчиками, а не кормовыми фуражирами. Поскольку сбор смолы может различаться в течение всего сезона, сбор данных был ограничен двумя неделями до заражения и двумя неделями после заражения. Наблюдая за количеством смоляных фуражиров, возвращающихся в колонию в течение определенного периода, мы смогли точно измерить изменения в сборе смолы за период до и после заражения. Другие методы измерения сбора смолы (например, накопление ее в улье) неточны, так как смола размещается по всей внутренней части гнезда в небольших трещинах и щелях и смешивается с различным количеством воска.
Анализ данных
Для описания изменений в снабжении смолой и пыльцой в течение двух периодов времени в наиболее простом варианте была рассчитана разница между общим числом фуражиров до и после заражения (сумма после минус сумма до) для каждой семьи. Поскольку данные имели нормальное распределение, был использован метод ANOVA для определения существенных различий (JMP V.9.0) в течение каждого года. Затем были объединены данные для незараженных семей и для зараженных аскосферозом в течение нескольких лет и проанализированы с использованием двусторонней ANOVA с такими основными параметрами, как год и заражение аскосферозом. Для данных 2008 года также были изучены различия в паразитарной нагрузке с использованием двусторонней ANOVA с такими основными параметрами, как уровень сбора смол и зараженность аскосферозом.
Во все годы размер выборки включал семьи, в которых смоляные фуражиры наблюдались как «до», так и «после» заражения. Семьи, имевшие клинические симптомы аскосфероза или другой инфекции (например, вируса деформации крыла) в период до заражения, в анализ не включались. В общей сложности из рассмотрения было удалено 5 семей (4 в 2008 году, 2 из каждой обработки и 1 семья с заражением американским гнильцом в 2009 году). Поскольку продолжительность наблюдений составляла примерно 1,5 часа на каждую семью и, следовательно, примерно 100 часов полевых наблюдений в течение всего эксперимента, несколько колоний с нулевыми фуражирами в один момент времени, вероятно, были либо результатом пропущенной выборки, либо другой причиной. Это привело к следующим размерам выборки: 2008 год
n = 7 незараженных и n = 10 аскосферозных; 2009 год-n = 8 незараженных колоний N = 8 американский гнилец, n = 8 аскосферозных и n = 9 Metarhizium; 2010 год-n = 6 незараженных и n = 7 аскосферозных.
Благодарности.
Мы благодарим г. Рейтера и Б. Ранума за помощь в поддержании колонии и сборе данных, а также М. Гоблирша, Дж.Гарднера, К. Ли и Р. Борбату за помощь в сборе данных; М. би, г. Хеймпеля и Дж. Эванса за первоначальные комментарии к этой рукописи; и К. Арельяно и Р. Муна за статистическую поддержку.
Примечания.
Конкурирующие интересы: авторы заявили, что никаких конкурирующих интересов не существует.
Финансирование: это исследование было поддержано грантом Национального научного фонда (IOS-0717530), присужденным MS, и стипендией аспиранта NSF, присужденной MSF. Спонсоры не играли никакой роли в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Перевод: Екатерина Егорова