
Автор: Джон Макмаллан
Перевод на русский язык: Елена Арифулина
Версия на английском языке (pdf)
Вступление
В Северном графстве Дублин, Ирландия, регионе, который имеет относительно автономный режим пчеловодства, с незначительным объёмом ввозимых в регион пчелосемей или маток, постепенно развивается толерантность (взаимная адаптация) к клещу Варроа. Это происходит с 2010 года без использования каких-либо мер против клеща, а также без селекции пчёл или специального отбора пчелосемей. Регион ограничен Ирландским морем на востоке, графствами Мит и Килдэр на западе, рекой Лиффи и пригородами Дублина на юге (Рис.1). Он имеет богатую историю, связанную с эпохой викингов, и к 841 году нашей эры был частью скандинавского поселения в Дублине. Власть викингов была свернута после битвы при Клонтарфе в 1014 году, но многие фамилии и топонимы региона до сих пор имеют скандинавское происхождение. Это плодородная земля с относительно плоским рельефом около 40 км в длину и 25 км в ширину. Пчеловодство региона имеет долгую историю, и как предполагается, оно было введено монахом Св. Молагой, жившим в седьмом веке, который построил церковь Ланн Бичер (Церковь Пасечника) недалеко от Балбриггана в Северном графстве Дублин. На каменной арке, датированной 1689 годом, в расположенном рядом Бреморском замке изображен монах, держащий роевню в форме колокола с летящими в неё пчёлами (Рис.2).
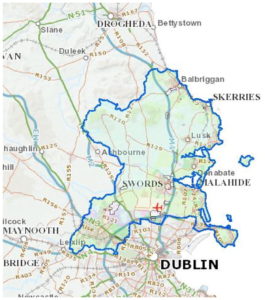
Рисунок 1.
Границы Северного графства Дублин выделены синим цветом. Регион ограничен Ирландским морем на востоке, графствами Мит и Килдэр на западе, рекой Лиффи и пригородами Дублина на юге.

Рисунок 2.
Каменная арка, датированная 1689 годом, в замке Бремор, Балбригган, примыкающем к церкви Св. Молаги в Северном графстве Дублин. На арке, по-видимому, изображен монах (справа), держащий роевню с летящими в неё пчёлами.
Нашествие Варроа
В регионах умеренного климата колонии пчёл обычно погибают через три-четыре года после первого заражения Варроа (Büchler, 1994). Начиная с ранней стадии заражения клещами Варроа в Западной Европе и Северной Америке были доступны методы лечения, которые были эффективны в борьбе с численностью клеща. Ежегодные однократные или более частые обработки пчелосемей теперь являются нормой в пчеловодстве, хотя уровень заражённости клещами пока стабильно высокий. Клеща Варроа и сопутствующие инфекции в настоящее время общепринято рассматривают как один из важных факторов гибели пчелосемей в мире (Rosenkranz, Aumeier, & Ziegelmann 2010). В Ирландии клещ Варроа впервые был обнаружен в 1998 году, и к сентябрю 2003 года он распространился на всю территорию Северного графства Дублин. Автор данной статьи выявил и опознал клеща на собственной пасеке, расположенной в центре графства, к северу от города Сордс (Рис.1), где он возник в результате появления по-соседству сильно заражённых пчелосемей, привезённых с инфицированного клещом юго-востока графства. Угроза для пчёл оказалась настолько незнакомой, что первоначальная реакция самой пострадавшей семьи состояла в том, что пчёлы построили толстую стенку из прополиса поперёк входа в улей, оставив только небольшой вход. Впоследствие заражённые семьи погибали, если не подвергались лечению, которое в нашем регионе обычно проводят с использованием химических препаратов на основе тимола.
Режим пчеловодства в регионе
Кочевое пчеловодство не распространено в этом в регионе, и в целом перемещение пчелосемей в регион или внутри него достаточно ограниченное. Исключение составляет малоинтенсивная интродукция местных ирландских пчеломаток Apis mellifera mellifera (пчела Galtee), размножаемых в графстве Типперэри, которая особенно активно велась в конце 1990-х и начале 2000-х годов. В последнее десятилетие в Северном графстве Дублин значительно увеличилось количество начинающих пчеловодов. Это произошло, по факту, во многом благодаря системе наставничества, в рамках которой опытные пчеловоды местной ассоциации (Fingal North Dublin BKA) курируют стартап-пасеки и снабжают их пчёлами из своих колоний (McMullan, 2012). Ассоциация приняла это решение в целях снижения распространения болезней, в частности американского гнильца, а также для ограничения ввоза в регион неместных пород Apis mellifera. Такая природная стационарная организация пчеловодства, когда плотность пасек невысока, а пчелосемьи самостоятельно выращивают себе замену, должна снижать вирулентность паразитов за счёт их вертикальной передачи, то есть передачи через потомство (Fries & Camazine, 2001). Это классический ответ «хозяев» на заражение иноземным паразитом при отсутствии внешнего лечения.
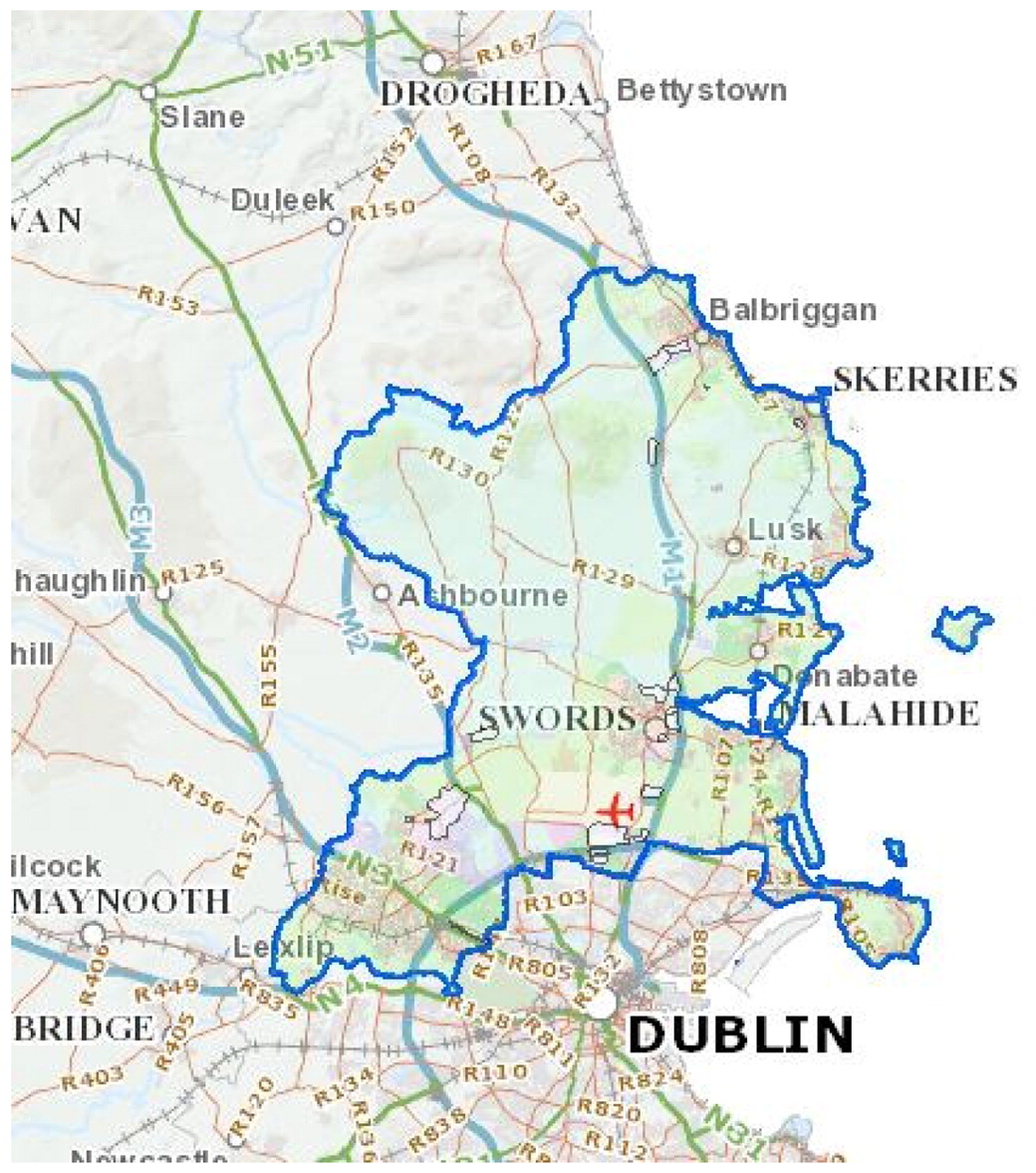
Рисунок 3. Схема взаимодействия паразита и хозяина. На диаграмме показано, как баланс (или толерантность) может возникнуть из-за изменчивости вирулентности паразита и восприимчивости хозяина (McMullan, 2012). При первом заражении пчелосемьи клещом Варроа (справа) семья обычно погибает из-за высокой вирулентности клеща и низкой устойчивости (высокой восприимчивости) медоносных пчел. Со временем, по мере естественного отбора и при отсутствии лечения вирулентность клеща снижается, а устойчивость пчёл повышается, вследствие чего возникает баланс отношений.
Демонстрация того, как это может произойти, приведена на рис.3. В своём существовании паразит зависит от хозяина, то есть он берёт ресурсы (питание) от хозяина, причем вирулентность отражает его способность нанести вред своему хозяину; с другой стороны, от восприимчивости хозяина зависит вероятность заражения паразитом. Согласно Шмид-Гемпелю и Коэллу (Schmid-Hempe, Koella, 1994), «изменчивость во взаимодействии хозяина и паразита оказывает значительное влияние на экологию и эволюцию паразитов, а также на эпидемиологию заболевания». Эта изменчивость создаёт возможности для адаптации. При низкой вирулентности паразита и низкой восприимчивости хозяина паразит не сможет поддерживать свое существование и погибнет. И наоборот, при высокой вирулентности и восприимчивости паразит может убить своего хозяина, вследствие чего погибают и хозяин, и паразит. Это обычный исход в ситуации, когда хозяина поражает паразит-экзот, с которым ранее хозяин не сталкивался.
К 2010 году в регионе стали проявляться более благоприятные отношения между клещами и медоносными пчелами. Как оказалось, пропуск ежегодной обработки от клеща, который чаще всего случался из-за неблагоприятных погодных условий в конце сезона, имел мало негативных последствий для пчелосемей. Уже более семи лет я не лечу своих пчёл, и всё меньше пчеловодов региона лечат пчёл в целях контроля клеща. К маю 2017 года почти две трети пчеловодов региона приняли подход, не основанный на лечении. Пчёлам обычно дают возможность выращивать своих собственных маток, как в случае родительского роя в роящейся пчелосемье, после того как пчеловод удаляет вторую семью со старой маткой, причём последняя затем остается в пределах региона. Значительное увеличение числа начинающих пчеловодов (с 2010 года количество членов выросло более чем в два раза) и политика снабжения новых пасек местными пчёлами привели к ощутимому росту числа пчелиных колоний, причём почти все они возникли внутри региона, без ввоза пчёл извне. По сути, паразиты (клещи и болезни, которые они передают) и пчелиные хозяева в их сложных взаимоотношениях были предоставлены самим себе, в попытках найти свой собственный эволюционный путь к взаимной терпимости. (Blacquière & Panziera, 2018). Эти условия похожи на ситуацию, в которой оказались дикие медоносные пчелы: в результате возникло множество диких колоний, заражённых клещом, которые выживают без какого-либо лечения (Fries, Imdorf, & Rosenkranz, 2006; Le Conte et al., 2007; Locke, 2016; Seeley, 2007).
Перемены в колониях медоносных пчёл
Пчеловоды региона наблюдают изменения, которые происходят в популяции медоносных пчёл с начала появления клеща Варроа. Первое время обычным явлением были признаки вируса деформации крыла (ВДК), которые обнаруживались во время осмотра ульев в конце лета: группы пчёл, повреждённых этим вирусом, ползающие по земле перед ульями. Со временем случаи ВДК пошли на убыль, и сегодня он стал редким явлением. За этот период в колониях пчёл Северного графства Дублин наблюдались и другие перемены: больше трутней и трутневого расплода размещаются ближе к центру пчелиного гнезда, зимняя смертность пчелосемей, согласно данным COLOSS для Северного графства Дублина, снизилась в период 2014 – 2017 гг. с 15,8 до 11,2%. Уровень смертности в Ирландии в целом в 2015/16 году составил 29,5% (Brodschneider et al., 2016). Во всем мире первичное заражение клещом Варроа обычно вело к быстрому, иногда резкому, росту популяции клещей и гибели пчелосемьи (Büchler, 1994). При толерантных или сбалансированных отношениях между клещом и пчелой-хозяином можно ожидать, что, хотя численность клеща колеблется в течение сезона, она будет одинаковой в одно и то же время каждый год. В ходе развития толерантных отношений хозяева и паразиты могут претерпевать изменения в биологии и поведении. Повреждение клеща пчёлами может способствовать развитию толерантных отношений. Как было показано, на идиосоме (основная часть тела клеща – прим. переводчика) могут появляться одна или две правильные симметричные дорсальные (спинные) ямки, которые можно рассматривать как «дефекты», а не как «повреждения», вызванные пчёлами (Davis, 2009). Исследование Lodesani, Vecchi, Tommasini и Bigliardi (1996) показало, что наличие дорсальных ямок у самок-основательниц приводило к снижению численности женского потомства: отсутствие дочерей первого поколения достигало 35,5% — по сравнению с 14,5% у самок без дорсальных ямок. Исследование Лодесани (Lodesani, 1996) также выявило, что распространённость этого дефекта у клещей в ячейках расплода и у клещей, упавших на дно улья, не отличается. Изменения продолжительности периода развития личинки пчелы в запечатанной ячейке может повлиять на время, доступное клещам для развития в ячейках расплода, что, соответственно, имеет решающее значение для воспроизводства женского потомства клеща (Büchler & Drescher, 1990). Кроме того, продолжительность стадии окукливания будет зависеть от температуры гнезда, и в этой связи было показано, что в ответ на угрозу паразита или патогена пчелосемья может повысить температуру (Hou, Li, Deng, & Diao, 2016; Starks, Blackie, & Seeley, 2000).
Исследования 2016-2017 годов
В 2016 и 2017 годах было проведено исследование, его цель – улучшить наше понимание динамики изменений, произошедших в Северном графстве Дублин за 14 лет с момента появления клеща Варроа. Эти изменения могут помочь выявить возможные причины развития толерантности к клещу, которую демонстрируют пчёлы. Исследование проводилось с апреля 2016 по май 2017 года, в центре графства на пасеке автора, состоящей из пяти ульев с пчёлами A. m. mellifera, которые с осени 2010 года не подвергались акарицидному или иному лечению. Все ульи имели цельное дно и сетку между полом и гнездовым отделением, то есть улья были без «открытого сетчатого дна». Ульи имели т.н. «модифицированную коммерческую» конструкцию с одним гнездовым корпусом, который по размеру был на 20% больше, чем в улье Лангстрота.
В апреле-мае 2016 года семьи быстро нарастили силу и имели в среднем по 8 рамок расплода в 11-ти рамочных гнездовых корпусах. Главный медосбор пришёлся на период с апреля до середины июня, от цветения весенних первоцветов до цветения древесных пород. В июле-августе наблюдался период падения взятка, который привёл к резкому сокращению количества расплода. Позже, с середины сентября по середину ноября 2016 года хороший взяток, в основном с плюща обыкновенного (Hedera helix), способствовал появлению сильных популяций «зимней» пчелы. В мае-июне 2016 года три семьи отстроили роевые маточники, у них были заменены пчеломатки, а отводки со старыми матками были перемещены на пасеки по-соседству. Все семьи успешно перезимовали, имея запасы более 20 кг, преимущественно плющевого мёда, из которых с ноября до конца марта обычно съедается менее 10 кг, так как A. m. mellifera отличается экономным потреблением. В апреле-мае 2017 года семьи вновь быстро выросли, имея в среднем более 8 рамок расплода в 11-ти рамочных гнездовых корпусах. К середине мая две семьи отстроили маточники и были в процессе замены пчеломатки.
В ходе исследования были изучены: сезонные уровни численности клещей; наличие повреждений клещей пчёлами; изменения дорсальных ямок на идиосоме клеща; распространённость клинических признаков вируса деформации крыла и статус вируса на основе метода ПЦР-РВ (полимеразная цепная реакция с обратной транскрипцией); изменение температуры гнездового отделения улья.
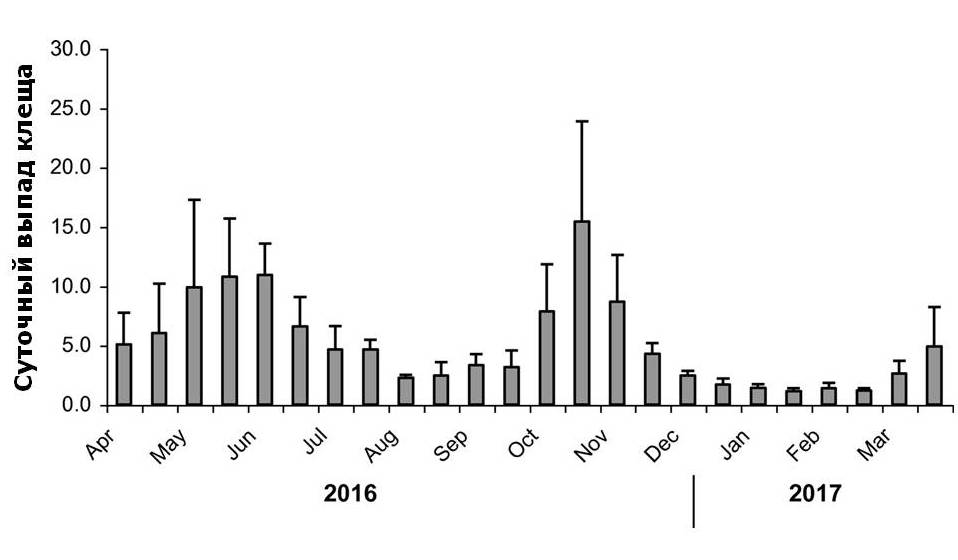
Сезонный выпад клеща
Для количественной оценки численности популяции клеща в пчелосемьях ведут подсчёт количества клещей, естественным образом осыпавшихся на дно улья. (Branco, Kidd, & Pickard, 2006). Клещи падают сквозь сетчатый экран на вкладыш, установленный поверх цельного дня улья. За двенадцать месяцев в период с апреля 2016 по март 2017 года осыпавшихся клещей собирали в каждом улье дважды в неделю. Полученные данные затем были сведены в график с разбивкой на двухнедельные периоды (Рис.4). График показывает, что заклещённость ульев растёт весной и осенью и падает в течение лета и зимы. Среднее количество осыпавшихся клещей в начале и конце двенадцатимесячного периода было одинаковым и составляло ок. 5шт. в день.
Повреждения клещей
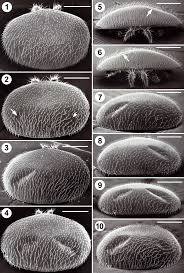
Для выявления возможного механизма, способствующего выработке толерантности, осыпавшихся клещей изучили на наличие повреждений. В течение десяти дней в октябре 2016 года мы ежедневно собирали и изучали опавших на дно ульев клещей. Поскольку на полу ульев не было муравьев или других крупных членистоногих, любые повреждения клещей, кроме правильных дорсальных ямок, были сделаны пчёлами. Было подсчитано количество живых и мёртвых клещей, а по состоянию их разделили на «неповреждённые», «повреждённые идиосомы» и «повреждённые конечности». Клещи с одной или двумя правильными дорсальными ямками не считаются повреждёнными (Davis, 2009). Большинство клещей, естественным образом осыпавшихся на дно улья, были мертвы, а повреждения наблюдались у 44% из них, включая 41% с повреждениями ног и 3% с неправильными повреждениями идиосомы (Рис.5). Поскольку ни муравьев, ни крупных членистоногих в ульях не было, этот вред клещам нанесли пчёлы. Повреждения конечностей у живых клещей были редкостью и были замечены только у 8 уже умирающих клещей из общего количества 1084. Количество мёртвых клещей, возможно, было завышено, так как некоторые из них могли быть живы ранее в тот день, когда они упали на вкладыш на дне улья.
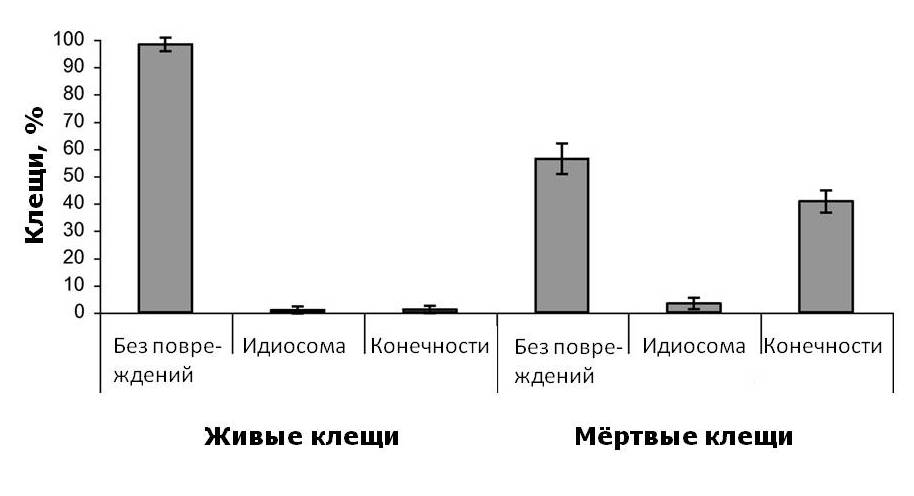
Периметр вкладыша на дне улья был смазан тонким слоем вазелина, прилипшие к мази клещи также были подсчитаны. Эта мера позволила учесть клещей, которые потенциально могли покинуть дно улья и вернуться в гнездовой корпус. К вазелину прилипли в общей сложности 36 клещей из 1084 (3,3%), что свидетельствует о слабом перемещении клещей к краям вкладыша. Кроме того, мы редко наблюдали клещей на дне улья, которые хоть как-то двигались.
Дорсальные ямки на идиосоме клещей
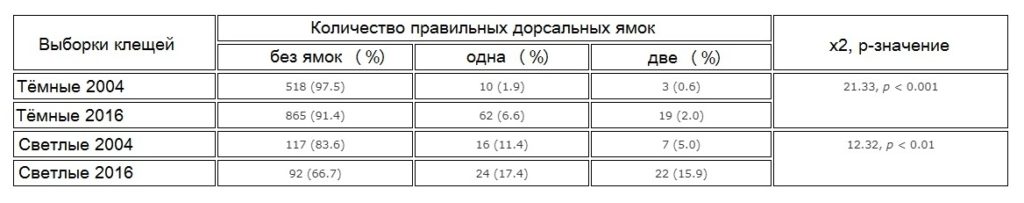
В ходе исследования клещей, упавших на дно ульев за десять дней в октябре 2016 года, также была изучена распространённость правильных дорсальных ямок (одиночных или двойных) на идиосоме насекомых. Были подсчитаны клещи с одной, двумя дорсальными ямками и без них, далее их разделили на самок-основательниц, отличающихся тёмной окраской, и светлоокрашенных дочерних самок. Кроме того, чтобы выявить какие-либо изменения в распространённости дорсальных ямок с течением времени, была изучена архивная выборка клещей, собранная во время осенней обработки против Варроа в 2004 году, вскоре после того, как клещи заразили пасеку. Клещей этой выборки сгруппировали аналогичным образом.
Распространённость дорсальных ямок была количественно оценена у взрослых (тёмноокрашенных) и дочерних (светлоокрашенных) клещей-самок для выборок октября 2016г. и 2004г. (табл. 1). Примечательно, что образец 2004г. содержал экземпляры пчелиных вшей (Braula coeca), которые с тех пор исчезли, возможно, в результате химической обработки. Распространённость дорсальных ямок в 2016г. оказалась значительно выше: в 3,4 раза у взрослых самок (χ2 = 21,33, p < 0,001) и в 2,0 раза у дочерних самок (χ2 = 12,32, p < 0,01).
Распространенность вируса деформации крыла
2 апреля 2017 года в каждом из пяти ульев выбрали по одной рамке расплода, имевших наибольшую площадь запечатанных ячеек расплода разного возраста. С рамок удалили взрослых пчёл, после чего каждую рамку поместили в отдельную клетку из нержавеющей перфорированной стали и перенесли в инкубатор с температурой 34,5°С и относительной влажностью воздуха 50-60%. Каждые восемь часов клетки переворачивали на новый бок, чтобы снизить различие температур в разных частях рамки расплода. Вылупившихся молодых пчёл собирали каждые восемь часов, изучали на наличие признаков деформации крыла, подсчитывали, упаковывали в пакеты и замораживали. Этот процесс продолжали, пока не вылупилась вся молодь. Молодых пчёл из всех пяти рамок затем протестировали на вирусный статус с помощью метода ПЦР-РВ (полимеразная цепная реакция с обратной транскрипцией), выполнив 40 циклов. Аналогичным образом изучили в полевых условиях взрослых летающих пчёл из каждого из пяти ульев.
Деформация крыла обнаружилась только у 24 из 7552 (0,32%) молодых пчёл, и ни в одном случае не было зафиксировано полностью усохших крыльев. Все образцы пчелиной молоди дали отрицательный результат теста на вирус деформации крыла. Два экземпляра из взрослых летающих пчёл показали положительный результат, и они происходили из тех же двух ульев, чьи рамки расплода имели наибольшую распространённость клинических признаков вируса у молодых пчёл.
Температура гнезда
В апреле 2017 года в течение десяти дней подряд с помощью датчиков, вставленных между сотами, в ульях измеряли температуру в центре гнезда (макс/мин). Для оценки влияния открытых сетчатых полов на температуру улья датчики разместили также под гнездом, над гнездом и по центру цельного дна улья, кроме того, измеряли температуру окружающей среды вне улья. Показания этих датчиков во всех пяти ульях записывали в течение четырёх дней подряд в апреле 2017 года.
Среднесуточная медианная температура во всех ульях за десятидневный период составила 35,2°C (± 0,08, n = 5). Среднесуточная медианная температура в центре цельных полов в ульях равнялась 28,2°C (± 1,4, n = 5). Соответствующая среднесуточная медианная температура окружающей среды составила 9,1°C (± 1,2, n = 5).
Выводы
Естественный выпад клеща во всех ульях продолжается с высокой скоростью и динамикой, соответствующей сезонной динамике пчелиного расплода (Fries, Camazine, & Sneyd, 1994), причём выпад клеща в начале и конце двенадцатимесячного периода остаётся одинаковым (Рис.4). Примечательно, что пчёлы демонстрируют толерантность на фоне по-прежнему высокого уровня выпада клеща. Между тем, согласно Морицу (Moritz, 1981), ежедневная небольшая потеря ресурсов, вызванная присутствием клеща, по всей видимости, мало влияет на здоровую пчелу. Как было обнаружено, уровни выпада клещей схожи в толерантных и неустойчивых к клещу диких колониях пчёл в лесу Арнот, США: в обоих случаях динамика роста заклещённости колоний была похожей (Seeley, 2007). Высокие показатели заклещённости были зарегистрированы на северо-востоке Бразилии в островной популяции пчёл, которая выживает с Варроа уже более тридцати лет (De Mattos, De Jong, & Soares, 2016).
Влияние других паразитов, в первую очередь, возбудителей вируса деформации крыла (ВДК), по-видимому, ослабевает. Зарегистрированная заболеваемость этим вирусом была высокой, когда клещ Варроа впервые появился в регионе, и оставалась высокой даже в первые годы, когда проводилось лечение пчёл. В первое время после отказа от лечения симптомы болезни проявлялись часто, в конце лета перед ульями нередко наблюдались мёртвые и умирающие пчёлы. Сегодня проявления ВДК встречаются нечасто. Относительно сильная заклещённость пчёл не привела к высокому уровню вирулентности вируса. Это соответствует результатами исследования Gisder, Aumeier и Genersch (2009), согласно которым само по себе заражение клещом не имеет прямой взаимосвязи с наличием признаков деформированных крыльев в колониях. Недавнее исследование (Brettell et al., 2017) показало, что единственным несомненным фактором повреждения крыльев у пчёл является высокий уровень вирусной нагрузки. Метод РТ-ПЦР продемонстировал, что пчёлы с повреждёнными крыльями имели самую высокую концентрацию возбудителей вируса, что также согласуется с результатами нашего исследования.
Большинство клещей, упавших на дно ульев, оказались мертвы. 44% упавших мёртвых клещей имели повреждения, включая 41% с повреждениями конечностей и 3% с неправильными повреждениями идиосомы (Рис.5). Повреждения конечностей у живых клещей были редкостью и были замечены только у нескольких умирающих клещей (у 8 из 1,084). По всей видимости, повреждение конечностей смертельно для клещей. Повреждения конечностей у мертвых клещей варьировались в широких пределах от оторванных кончиков (коготков) одной лапки до полностью отсутствующих нескольких конечностей. В свою очередь правильные повреждения (дорсальные ямки) на идиосоме клещей стали гораздо более распространёнными по сравнению с 2004г.: в 3,5 и 2,0 раза у взрослых самок-основательниц и дочерних самок соответственно. По данным Лодесани (Lodesani, 1996), это свидетельствует о значительном снижении плодовитости клещей, что должно способствовать выработке толерантности к Варроа.
Не обнаружено никаких признаков повышения температуры пчелиного гнезда, по сравнению с периодом до нашествия Варроа. Это наблюдение совпадает с выводами Левина и Коллисона (Levin, Collison, 1990). Таким образом, нет никаких свидетельств того, что в результате появления Варроа и повышения температуры гнезда продолжительность развития личинок пчёл в запечатанных ячейках уменьшилась. Ле Конте и Арнольд (Le Conte, Arnold, 1988) показали, что клещ Варроа предпочитает ячейки с температурой ниже 33°C и поэтому чаще выбирает ячейки с трутневым расплодом, который традиционно размещается по краю рамки расплода (Winston, 1987). Между тем всё чаще в Северном графстве Дублин трутневый расплод размещается выше и ближе к центру гнезда, в местах с более высокой температурой. Ожидаемое сокращение периода развития личинок трутней в запечатанных ячейках и, соответственно, периода размножения клеща, по-видимому, может стать примером работы естественного отбора, призванного ускорить выработку толерантности к варроа (Jandricic & Otis, 2003).
Аналогичным образом, использование цельного дна в ульях, как в данном исследовании, также может способствовать выработке толерантности. Как было показано, лишь небольшая часть упавших на дно улья клещей (3,3%) переместилась к краям дна и потенциально могла вернуться в гнездо. Кроме того, Коффи (Coffey, 2007) продемонстрировал, что в регионах умеренного климата первоначальное сокращение роста популяции клещей было менее устойчивым при использовании открытого сетчатого дна в ульях. Весной между центром цельного пола и внешней средой может возникать большая разница температур (в нашем исследовании она достигала 19°C). При наличии открытого сетчатого дна температура в этой точке улья будет стремиться к температуре окружающей среды, снижая температуру до 19°C. Следовательно, использование открытых сетчатых полов в регионах умеренного климата подвергает пчелосемьи большему риску в критически важный весенний период, когда семьи наращивают силу, и тем самым даёт клещам Варроа потенциальное репродуктивное преимущество.
Автор: Джон Макмаллан, бывший научный сотрудник Тринити-Колледжа, Дублин, Ирландия